by Dr. Boris Zakharov
ABOUT THE AUTHOR

Dr. Boris Zakharov
Professor Boris Zakharov began his professional career in the “Kedrovaya Pad Natural Reserve” (Far East State University, Vladivostok, Russia) as a research scientist, where he participated in and performed various scientific projects in field zoology. Dr. Zakharov's Doctoral research was on horse flies (Diptera, Tabanidae), beginning in 1983 at Novosibirsk Biological Institute. The results of this study were published in his papers and concern systematics, ecology, biogeography and cattle protection from blood sucking diptera in Siberia.
After his arrival in the United States, Dr. Zakharov participated in a project on ground spiders of Australia and New Zealand in the American Museum of Natural History, where he has worked from 1996 to 2005 as a Curatorial Assistant in the Spider laboratory in a Department of Invertebrate Zoology. Currently, he occupies the position of a visiting scientist at the American Museum of Natural History, where he continues the study of ground spiders of the world.
Professor Boris Zakharov began to teach in 2005 at Hostos Community College/CUNY. Boris joined the Natural Sciences department at LaGuardia in 2009.
Trichobothria are mechanoreceptors located on the dorsal surface of tarsus, metatarsus, and tibia of walking legs and pedipalps of spiders. They are thin hairs emerged from a cup shaped sockets sensitive to minute air movement.
Trichobothria are cuticle mechanoreceptors highly sensitive to small air movement. Dahl (1883) first suggested that these structures are auditory mechanoreceptors and named them Hörhaare. Later, he introduced the term trichobothria (Dahl, 1911). Further studies (Palmgren, 1936; Pauly, 1956; Schömann, 1956; Hoffmann, 1965) show that trichobothria respond to airborne stimuli such as gas particle displacement and have to be regarded not as sound detectors, but rather as “touch-in-distance” mechanoreceptors. Electrophysiological studies (Görner, 1965; Christian 1971, 1972; Barth, 2002) support this conclusion and suggest a specific mechanism of sensory input transduction.
Trichobothria are located on the tarsus, metatarsus, and tibia of spider legs. As a rule, they are organized in groups or rows, especially on the dorsal leg surface. The large wandering spider Cupienius has almost 1,000 trichobothria: about 100 on each leg and about 50 on each palp.

Ctenidae: Cupienius coccineus (female), Talamanca Reserve San Gerardo, Rivas, San José, Costa Rica. Photo credit: Tom Murray/Flickr.
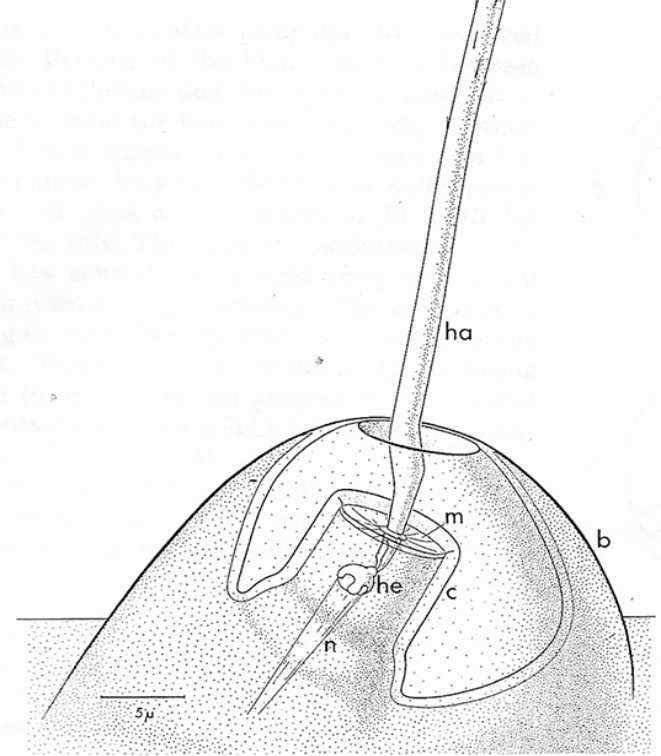
In gnaphosid spiders trichobothria are arranged in 1-3 rows: one dorsal row presents in all ground spiders and one or two lateral rows from both sides. As a rule, web spiders have much fewer trichobothria than ground spiders. This difference demonstrates higher importance of these sensory organs for ground wandering hunters than web spiders who more rely on information they receive from web vibrations. The arrangement of trichobothria is species specific and often is used for spiders’ classification and study of their evolutionary relationships. Trichobothrium has two components: 1) long thin hair or trichos and 2) cup shaped cuticle socket or bothrium. The ground plan of trichobothria construction in general is similar among spiders (Görner, 1965; Barth, 2002; Haupt, 2003; Felix, 2010) and other arachnids (Foelix&Schabronath, 1983; Haupt, 1996). It has the bowl-shaped socket in the cuticle with a single long hollow inside filament at the center (see Fig. 1).
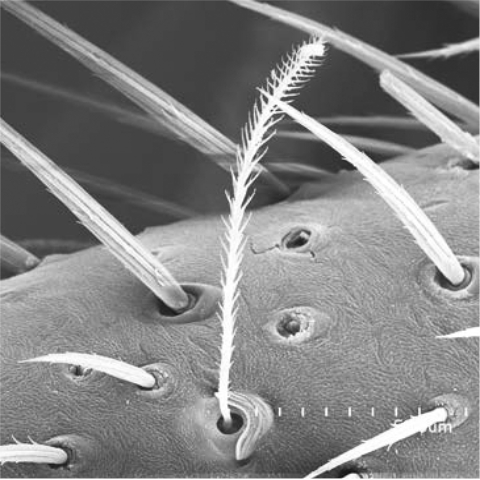
Figure 1. Trichobothria of Drassylus aprinus (Banks, 1901).
Typical hair length of this organ vary from 100 μm up to 1400 μm. The hair shaft of trichobothrium has a feathery-looking surface curved distally. The feathery surface of hair increases drag forces and thus creates a high mechanical sensitivity to air movement. The plane of the curvature is inclined from the leg’s main long axis at an angle of 5° to 30°. Depth and diameter of the sockets increase with the length of the hair. However, the angle through which the hair can be deflected between the two edges of the socket remains constant at 25°- 35°, because the socket of a long hair is not only broader but also deeper. Stretched across the bottom of the socket is a cuticular membrane that bears the hair shaft.
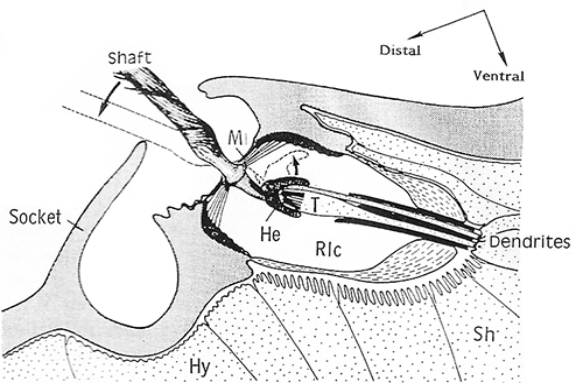
The length of the hairs increases distally on every limb section. All hairs have similar shape and are lightly inclined distally towards the tip of the leg (see Fig. 3). The socket is surrounded by distal and proximal plates that may have distal and proximal margins. Shape of the plates varies among spiders and often have more or less high ridges.
Proximally, hair rests inside a convex cap or helmet associated with dendrites of sensory neurons. Air movement pressing on the hair of the trichobothrium, bends it. The hair deflection energy is converted in sensory cells into the action potential. The sensitivity of trichobothria is amazing. The long hair shaft is suspended in a very thin (0.5 μm) cuticular membrane. Even a very light air current of around 1 mm/sec makes it quiver. An energy less than a quantum of green light is sufficient to excite the sensory cell. Different deflection directions stimulate different sensory cells, which number vary from 3 to 11. This creates effect of ability to monitor air movement from different directions. Each sensory cell reacts to displacements within a particular sector with a light overlapping among sectors.
The trichobothria receptors have distinct frequency tuning, with best frequencies between 50 and 120 Hz, which perfectly fit to the buzz produced by a flying nearby fly. The lack of spontaneous activity and marked phasic nature of the responses to trichobothria, which react mainly to the speed with which they are deflected, make these sensilla ideally suited to monitor the typically very inconstant stimuli. The irregular noise does not stimulate these sensilla, whereas a sinusoidal stimulus at 100 Hz is responded to in a few seconds.
Trichobothria allow spider not only to sense presence of the fly at distance of as much as 30 cm, but to identify its movement speed and direction to make a successful jump even in the absolute dark or when the spider is blinded. Usually this drama lasts less than 1.5 sec. The spider uses trichobothria not only to detect prey but also as a warning system against predatory wasps (Palpimonidae) and parasitic Neuroptera (Mantispidae).
References
Barth FG 2002 A spider’s world. Senses and behavior. Springer Berlin, Heidelberg, New York, Barcelona, Hong Kong, London, Milan, Paris, Tokyo.
Christian UH 1971 Zur Feinstruktur der Trichobothrien der Winkelspinne Tegenaria derhami (Scopoli) (Agelenidae, Araneae). Cytologie 4: 172-185.
Christian UH 1972 Trichobothrien ein Mechanorezeptor bei Spinnen. Elektronenmikroscopische Befunde bei der Winkelspinne Tegenaria derhami (Scopoli) (Agelenidae, Araneae). Verh. Dtsch. Zool. Ges. 66: 31-36.
Dahl F 1883 Über die Hörhaare bei den Arachniden. Zool. Anz. 6: 267-270.
Dahl F 1911 Die Hörhaare (Trichobothrien) und das System der Spinnentiere. Zool. Anz. 37: 522-532.
Foelix RF & Schabronath J 1983 The fine structure of scorpions sensory organs. I. Tarsal sensilla Bulletin of the British Arachnological Society 6: 53-67.
Foelix RF 2010 Biology of spiders. OUP USA/Georg Thieme Verlag, 330 pp.
Görner P 1965 Mehrfach innervierte Mechanorezeptoren bei Spinnen. Naturwissenschaften 52: 437
Görner P & Andrews P 1969 Trichobothrien, ein Ferntastsinnesorgan bei Webspinnen (Araneen). Z. vergl. Physiol. 64: 301-317.
Haupt J 2003 The Mesothelae — a monograph of an exceptional group of spiders (Araneae: Mesothelae) (Morphology, behavior, ecology, taxonomy, distribution and phylogeny). Zoologica. Originalabhandlungen aus dem Gesamtgebiet der Zoologie 154: 26 pp.
Palmgren P 1936 Experimentelle Untersuchungen über die Funktion der Trichobothrien bei Tegenaria derhami. Scop. Acta Zool. Fenn, 19: 3-27.
Schömann K 1956 Zur Biologie van Polyxenus lagurus (L. 1758). Zool. Jb. Abt. Systemat. 84: 195-256.